
DUSP-1 Induced by PGE2 and PGE1 Attenuates IL-1β-Activated MAPK Signaling, Leading to Suppression of NGF Expression in Human Intervertebral Disc Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
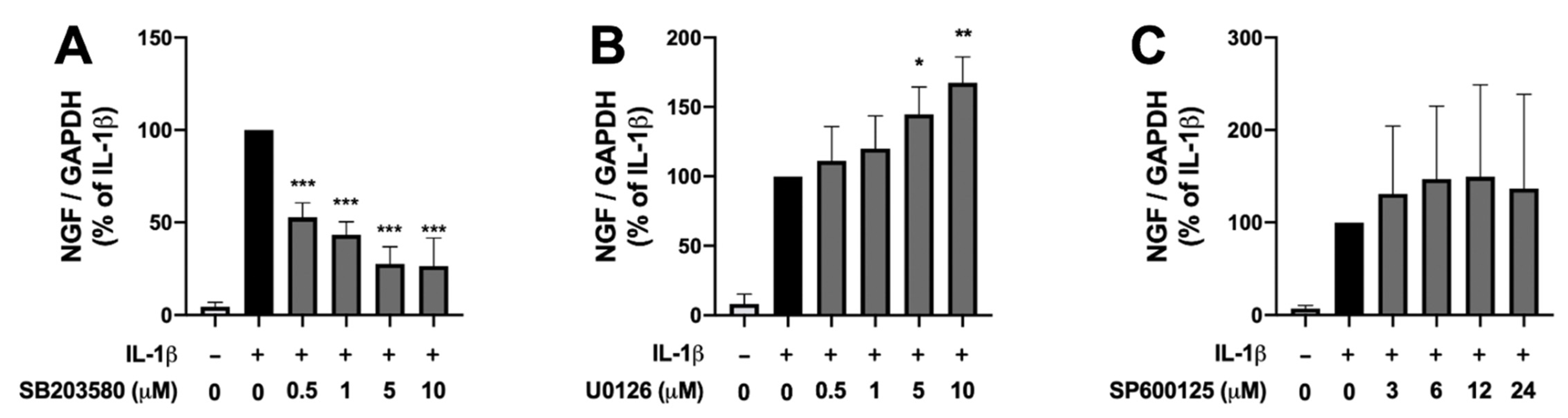
2.1. Effects of Inhibitors of Various MAPK Pathways on IL-1β-Induced NGF Expression in Human IVD Cells
2.2. PGE2 and PGE1 Suppressed IL-1β-Induced Phosphorylation of MAPKs in Human IVD Cells
2.3. PGE2 and PGE1 Enhanced DUSP-1 Expression in Human IVD Cells
2.4. DUSP-1 Knockdown by Small Interfering RNA (siRNA) Transfection in Human IVD Cells
2.5. IL-1β-Induced Phosphorylation of MAPKs Was Enhanced and Prolonged in DUSP-1 Knockdown Cells
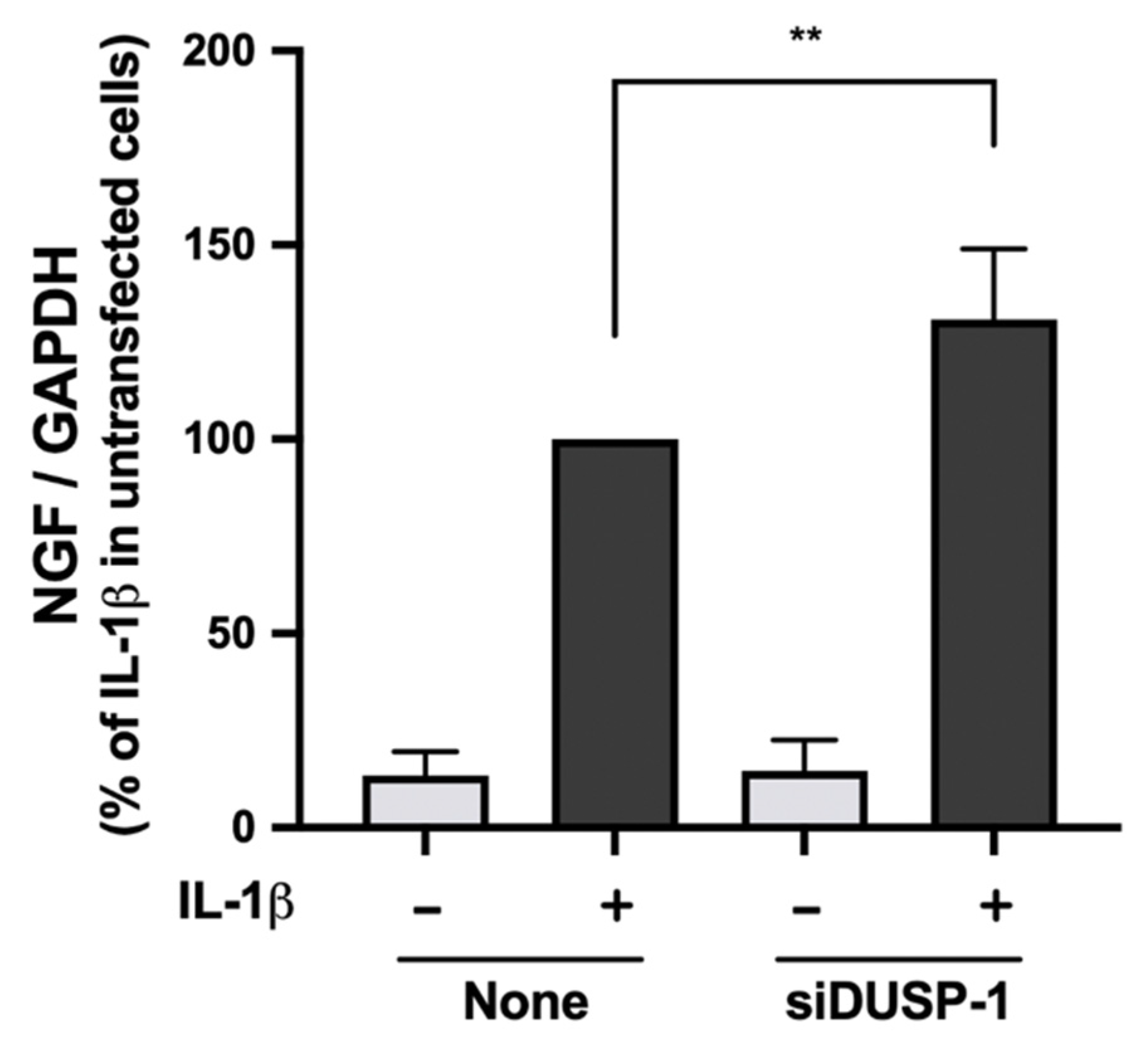
2.6. IL-1β-Induced NGF Expression Was Enhanced in DUSP-1 Knockdown Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Western Blotting
4.3. Real-Time Reverse Transcription–Polymerase Chain Reaction
4.4. siRNA Transfection
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vos, T.; Abajobir, A.A.; Abbafati, C.; Abbas, K.M.; Abate, K.H.; Abd-Allah, H.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; Aboyans, V.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Aoki, Y.; Ohtori, S. Resolving discogenic pain. Eur. Spine J. 2008, 17 (Suppl. 4), 428–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzer, A.C.; Aprill, C.N.; Derby, R.; Fortin, J.; Kine, G.; Bogduk, N. The prevalence and clinical features of internal disc disruption in patients with chronic low back pain. Spine 1995, 20, 1878–1883. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Roberts, S.; Smith, S.; Menage, J.; Ghosh, P. Increased nerve and blood vessel ingrowth associated with proteoglycan depletion in an ovine anular lesion model of experimental disc degeneration. Spine 2002, 27, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Snow, D.M.; Lemmon, V.; Carrino, D.A.; Caplan, A.I.; Silver, J. Sulfated proteoglycans in astroglial barriers inhibit neurite outgrowth in vitro. Exp. Neurol. 1990, 109, 111–130. [Google Scholar] [CrossRef]
- Shen, B.; Melrose, J.; Ghosh, P.; Taylor, F. Induction of matrix metalloproteinase-2 and -3 activity in ovine nucleus pulposus cells grown in three-dimensional agarose gel culture by interleukin-1β: A potential pathway of disc degeneration. Eur. Spine J. 2003, 12, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Melrose, J.; Shu, C.; Young, C.; Ho, R.; Smith, M.M.; Young, A.A.; Smith, S.S.; Gooden, B.; Dart, A.; Podadera, J.; et al. Mechanical destabilization induced by controlled annular incision of the intervertebral disc dysregulates metalloproteinase expression and induces disc degeneration. Spine 2012, 37, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Freemont, A.J.; Peacock, T.E.; Goupille, P.; Hoyland, J.A.; O’Brien, J.; Jayson, M.I. Nerve ingrowth into diseased intervertebral disc in chronic back pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef]
- Coppes, M.H.; Marani, E.; Thomeer, R.T.; Groen, G.J. Innervation of “painful” lumbar discs. Spine 1997, 22, 2342–2349. [Google Scholar] [CrossRef]
- Ozawa, T.; Ohtori, S.; Inoue, G.; Aoki, Y.; Moriya, H.; Takahashi, K. The degenerated lumbar intervertebral disc is innervated primarily by peptide-containing sensory nerve fibers in humans. Spine 2006, 31, 2418–2422. [Google Scholar] [CrossRef] [Green Version]
- Levi-Montalcini, R. The nerve growth factor 35 years later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Lewin, G.R.; Ritter, A.M.; Mendell, L.M. Nerve growth factor-induced hyperalgesia in the neonatal and adult rat. J. Neurosci. 1993, 13, 2136–2148. [Google Scholar] [CrossRef]
- Woolf, C.J.; Ma, Q.P.; Allchorne, A.; Poole, S. Peripheral cell types contributing to the hyperalgesic action of nerve growth factor in inflammation. J. Neurosci. 1996, 16, 2716–2723. [Google Scholar] [CrossRef]
- Dyck, P.J.; Peroutka, S.; Rask, C.; Burton, E.; Baker, M.K.; Lehman, K.A.; Gillen, D.A.; Hokanson, J.L.; O’Brien, P.C. Intradermal recombinant human nerve growth factor induces pressure allodynia and lowered heat-pain threshold in humans. Neurology 1997, 48, 501–505. [Google Scholar] [CrossRef]
- Svensson, P.; Cairns, B.E.; Wang, K.; Arendt-Nielsen, L. Injection of nerve growth factor into human masseter muscle evokes long-lasting mechanical allodynia and hyperalgesia. Pain 2003, 104, 241–247. [Google Scholar] [CrossRef]
- Ruiz, G.; Ceballos, D.; Baños, J.E. Behavioral and histological effects of endoneurial administration of nerve growth factor: Possible implications in neuropathic pain. Brain Res. 2004, 1011, 1–6. [Google Scholar] [CrossRef]
- Freemont, A.J.; Watkins, A.; Le Maitre, C.; Baird, P.; Jeziorska, M.; Knight, M.T.; Ross, E.R.; O’Brien, J.P.; Hoyland, J.A. Nerve growth factor expression and innervation of the painful intervertebral disc. J. Pathol. 2002, 197, 286–292. [Google Scholar] [CrossRef]
- Markman, J.D.; Bolash, R.B.; McAlindon, T.E.; Kivitz, A.J.; Pombo-Suarez, M.; Ohtori, S.; Roemer, F.W.; Li, D.J.; Viktrup, L.; Bramson, C.; et al. Tanezumab for chronic low back pain: A randomized, double-blind, placebo- and active-controlled, phase 3 study of efficacy and safety. Pain 2020, 161, 2068–2078. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Che, M.; Xin, J.; Zheng, Z.; Li, J.; Zhang, S. The role of IL-1β and TNF-α in intervertebral disc degeneration. Biomed. Pharm. 2020, 131, 110660. [Google Scholar] [CrossRef]
- Le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. The role of interleukin-1 in the pathogenesis of human intervertebral disc degeneration. Arthritis Res. Ther. 2005, 7, R732–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiler, C.; Nerlich, A.G.; Bachmeier, B.E.; Boos, N. Expression and distribution of tumor necrosis factor α in human lumbar intervertebral discs: A study in surgical specimen and autopsy controls. Spine 2005, 30, 44–53, discussion 54. [Google Scholar] [CrossRef]
- Burke, J.G.; RW, G.W.; Conhyea, D.; McCormack, D.; Dowling, F.E.; Walsh, M.G.; Fitzpatrick, J.M. Human nucleus pulposis can respond to a pro-inflammatory stimulus. Spine 2003, 28, 2685–2693. [Google Scholar] [CrossRef]
- Ohtori, S.; Suzuki, M.; Koshi, T.; Takaso, M.; Yamashita, M.; Inoue, G.; Yamauchi, K.; Orita, S.; Eguchi, Y.; Kuniyoshi, K.; et al. Proinflammatory cytokines in the cerebrospinal fluid of patients with lumbar radiculopathy. Eur. Spine J. 2011, 20, 942–946. [Google Scholar] [CrossRef] [Green Version]
- Ohtori, S.; Miyagi, M.; Eguchi, Y.; Inoue, G.; Orita, S.; Ochiai, N.; Kishida, S.; Kuniyoshi, K.; Nakamura, J.; Aoki, Y.; et al. Epidural administration of spinal nerves with the tumor necrosis factor-α inhibitor, etanercept, compared with dexamethasone for treatment of sciatica in patients with lumbar spinal stenosis: A prospective randomized study. Spine 2012, 37, 439–444. [Google Scholar] [CrossRef] [Green Version]
- Ohtori, S.; Miyagi, M.; Eguchi, Y.; Inoue, G.; Orita, S.; Ochiai, N.; Kishida, S.; Kuniyoshi, K.; Nakamura, J.; Aoki, Y.; et al. Efficacy of epidural administration of anti-interleukin-6 receptor antibody onto spinal nerve for treatment of sciatica. Eur. Spine J. 2012, 21, 2079–2084. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Kokubun, S. Changes with age in proteoglycan synthesis in cells cultured in vitro from the inner and outer rabbit annulus fibrosus. Responses to interleukin-1 and interleukin-1 receptor antagonist protein. Spine 2000, 25, 166–169. [Google Scholar] [CrossRef]
- Abe, Y.; Akeda, K.; An, H.S.; Aoki, Y.; Pichika, R.; Muehleman, C.; Kimura, T.; Masuda, K. Proinflammatory cytokines stimulate the expression of nerve growth factor by human intervertebral disc cells. Spine 2007, 32, 635–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Maitre, C.L.; Hoyland, J.A.; Freemont, A.J. Catabolic cytokine expression in degenerate and herniated human intervertebral discs: IL-1β and TNFα expression profile. Arthritis Res. Ther. 2007, 9, R77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purmessur, D.; Freemont, A.J.; Hoyland, J.A. Expression and regulation of neurotrophins in the nondegenerate and degenerate human intervertebral disc. Arthritis Res. Ther. 2008, 10, R99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelofs, P.D.; Deyo, R.A.; Koes, B.W.; Scholten, R.J.; van Tulder, M.W. Nonsteroidal anti-inflammatory drugs for low back pain: An updated Cochrane review. Spine 2008, 33, 1766–1774. [Google Scholar] [CrossRef] [PubMed]
- Alimasi, W.; Sawaji, Y.; Endo, K.; Yorifuji, M.; Suzuki, H.; Kosaka, T.; Shishido, T.; Yamamoto, K. Regulation of nerve growth factor by anti-inflammatory drugs, a steroid, and a selective cyclooxygenase 2 inhibitor in human intervertebral disc cells stimulated with interleukin-1. Spine 2013, 38, 1466–1472. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Sawaji, Y.; Alimasi, W.; Suzuki, H.; Endo, K.; Tanaka, H.; Yorifuji, M.; Kosaka, T.; Shishido, T.; Yamamoto, K. PGE1 Attenuates IL-1β-induced NGF Expression in Human Intervertebral Disc Cells. Spine 2016, 41, E710–E716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thalhamer, T.; McGrath, M.A.; Harnett, M.M. MAPKs and their relevance to arthritis and inflammation. Rheumatology 2008, 47, 409–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Karin, M. Mammalian MAP kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [Green Version]
- Rincón, M.; Davis, R.J. Regulation of the immune response by stress-activated protein kinases. Immunol. Rev. 2009, 228, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Keyse, S.M. Protein phosphatases and the regulation of mitogen-activated protein kinase signalling. Curr. Opin. Cell Biol. 2000, 12, 186–192. [Google Scholar] [CrossRef]
- Lawan, A.; Shi, H.; Gatzke, F.; Bennett, A.M. Diversity and specificity of the mitogen-activated protein kinase phosphatase-1 functions. Cell Mol. Life Sci. 2013, 70, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Wancket, L.M.; Frazier, W.J.; Liu, Y. Mitogen-activated protein kinase phosphatase (MKP)-1 in immunology, physiology, and disease. Life Sci. 2012, 90, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.Z.; Yun, Z.; Wang, W.; Ma, B.A. Dual specificity phosphatase 1 has a protective role in osteoarthritis fibroblastlike synoviocytes via inhibition of the MAPK signaling pathway. Mol. Med. Rep. 2017, 16, 8441–8447. [Google Scholar] [CrossRef]
- Salojin, K.V.; Owusu, I.B.; Millerchip, K.A.; Potter, M.; Platt, K.A.; Oravecz, T. Essential Role of MAPK Phosphatase-1 in the Negative Control of Innate Immune Responses. J. Immunol. 2006, 176, 1899–1907. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.; Han, J.; Hui, L. MAPK signaling in inflammation-associated cancer development. Protein Cell 2010, 1, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef]
- Zarubin, T.; Han, J. Activation and signaling of the p38 MAP kinase pathway. Cell Res. 2005, 15, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Studer, R.K.; Vo, N.V.; Sowa, G.A.; Kang, J.D. p38 MAPK inhibition selectively mitigates inflammatory mediators and VEGF production in AF cells co-cultured with activated macrophage-like THP-1 cells. Osteoarthr. Cartil. 2009, 17, 1662–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Studer, R.K.; Gilbertson, L.G.; Georgescu, H.; Sowa, G.; Vo, N.; Kang, J.D. p38 MAPK inhibition modulates rabbit nucleus pulposus cell response to IL-1. J. Orthop. Res. 2008, 26, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.R.; Sekiguchi, Y.; Kikuchi, S.; Scott, B.; Medicherla, S.; Protter, A.; Campana, W.M. Inhibition of p38 MAP kinase activity enhances axonal regeneration. Exp. Neurol. 2003, 184, 606–614. [Google Scholar] [CrossRef]
- Schäfers, M.; Svensson, C.I.; Sommer, C.; Sorkin, L.S. Tumor necrosis factor-α induces mechanical allodynia after spinal nerve ligation by activation of p38 MAPK in primary sensory neurons. J. Neurosci. 2003, 23, 2517–2521. [Google Scholar] [CrossRef] [Green Version]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy—From molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta 2005, 1754, 253–262. [Google Scholar] [CrossRef]
- Abraham, S.M.; Lawrence, T.; Kleiman, A.; Warden, P.; Medghalchi, M.; Tuckermann, J.; Saklatvala, J.; Clark, A.R. Antiinflammatory effects of dexamethasone are partly dependent on induction of dual specificity phosphatase 1. J. Exp. Med. 2006, 203, 1883–1889. [Google Scholar] [CrossRef] [Green Version]
- Abraham, S.M.; Clark, A.R. Dual-specificity phosphatase 1: A critical regulator of innate immune responses. Biochem. Soc. Trans. 2006, 34, 1018–1023. [Google Scholar] [CrossRef] [Green Version]
- Nieminen, R.; Korhonen, R.; Moilanen, T.; Clark, A.R.; Moilanen, E. Aurothiomalate inhibits cyclooxygenase 2, matrix metalloproteinase 3, and interleukin-6 expression in chondrocytes by increasing MAPK phosphatase 1 expression and decreasing p38 phosphorylation: MAPK phosphatase 1 as a novel target for antirheumatic drugs. Arthritis Rheum. 2010, 62, 1650–1659. [Google Scholar] [CrossRef]
- Hashizume, M.; Mihara, M. High molecular weight hyaluronic acid inhibits IL-6-induced MMP production from human chondrocytes by up-regulating the ERK inhibitor, MKP-1. Biochem. Biophys. Res. Commun. 2010, 403, 184–189. [Google Scholar] [CrossRef]
- Maekawa, A.; Sawaji, Y.; Endo, K.; Kusakabe, T.; Konishi, T.; Tateiwa, T.; Masaoka, T.; Shishido, T.; Yamamoto, K. Prostaglandin E2 induces dual-specificity phosphatase-1, thereby attenuating inflammatory genes expression in human osteoarthritic synovial fibroblasts. Prostaglandins Other Lipid Mediat. 2021, 154, 106550. [Google Scholar] [CrossRef]
- Kiriyama, M.; Ushikubi, F.; Kobayashi, T.; Hirata, M.; Sugimoto, Y.; Narumiya, S. Ligand binding specificities of the eight types and subtypes of the mouse prostanoid receptors expressed in Chinese hamster ovary cells. Br. J. Pharmacol. 1997, 122, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Onda, A.; Kikuchi, S.; Yabuki, S.; Otani, K.; Nikaido, T.; Watanabe, K.; Konno, S. Limaprost alfadex and nonsteroidal anti-inflammatory drugs for sciatica due to lumbar spinal stenosis. Eur. Spine J. 2013, 22, 794–801. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J.; Kobayashi, H.; Wakabayashi, S.; Deguchi, M.; Ito, H.; Mogami, Y.; Tanikawa, H.; Nakagawa, H.; Moriya, H.; Ashizawa, R.; et al. The effect of a prostaglandin E1 derivative on the symptoms and quality of life of patients with lumbar spinal stenosis. J. Orthop. Sci. 2013, 18, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Pfirrmann, C.W.; Metzdorf, A.; Zanetti, M.; Hodler, J.; Boos, N. Magnetic resonance classification of lumbar intervertebral disc degeneration. Spine 2001, 26, 1873–1878. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kusakabe, T.; Sawaji, Y.; Endo, K.; Suzuki, H.; Konishi, T.; Maekawa, A.; Murata, K.; Yamamoto, K. DUSP-1 Induced by PGE2 and PGE1 Attenuates IL-1β-Activated MAPK Signaling, Leading to Suppression of NGF Expression in Human Intervertebral Disc Cells. Int. J. Mol. Sci. 2022, 23, 371. https://doi.org/10.3390/ijms23010371
Kusakabe T, Sawaji Y, Endo K, Suzuki H, Konishi T, Maekawa A, Murata K, Yamamoto K. DUSP-1 Induced by PGE2 and PGE1 Attenuates IL-1β-Activated MAPK Signaling, Leading to Suppression of NGF Expression in Human Intervertebral Disc Cells. International Journal of Molecular Sciences. 2022; 23(1):371. https://doi.org/10.3390/ijms23010371
Chicago/Turabian StyleKusakabe, Takuya, Yasunobu Sawaji, Kenji Endo, Hidekazu Suzuki, Takamitsu Konishi, Asato Maekawa, Kazuma Murata, and Kengo Yamamoto. 2022. "DUSP-1 Induced by PGE2 and PGE1 Attenuates IL-1β-Activated MAPK Signaling, Leading to Suppression of NGF Expression in Human Intervertebral Disc Cells" International Journal of Molecular Sciences 23, no. 1: 371. https://doi.org/10.3390/ijms23010371